Self-Similarity and Fractal StructureThere is not just one timeline in the brain, there are many. We can arbitrarily construct such a timeline at any level of resolution, not just for the neural network as a whole but also for the neurons and synapses (even molecules) within it. What is the significance of a collection of time series relative to internal events? The importance of the time element becomes immediately obvious when we begin to consider Markov and non-Markov behavior in neural networks. But before getting into that, we can consider what happens when the extent of the mapping interval varies. 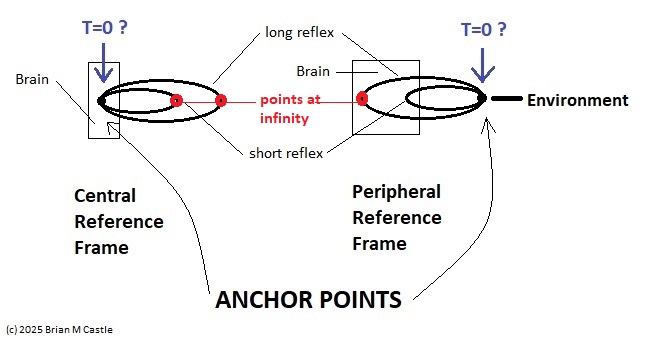 The figure defines a concept called "anchor", which we'll use in various ways during this discussion. Loosely speaking, it's related to where we put the electrode, which in turn defines our mapping origin T=0. From the standpoint of a black box (the organism) interacting with the environment, we can arbitarily define the origin to be the environmental interface, because we can easily measure it (at least, more easily than invading the black box in most cases). And, this definition also makes sense from the point of view of a central timeline, because the origin is then by definition the point of singularity. The Hawaiian EarringConsidering brain architecture in terms of feedback loops, it is clear that there are many loops with differing conduction delays. A monosynaptic reflex arc through the neck will have a shorter loop time than one through the toe. A molecular loop could be on the order of microseconds, while a circadian loop takes days. The various loop times result in differing radii after we compactify the associated timelines. In a very large network consisting of billions of neurons, we can imagine a rich loop structure, in such a way that for any given radius, we are likely to find an example of it - or at least an example near it. In other words, the loops tend to fill space, and since the timeline is fundamentally a place map, the loop construction “covers” the singularity at T=0, much as our visual perception covers the blind spot. 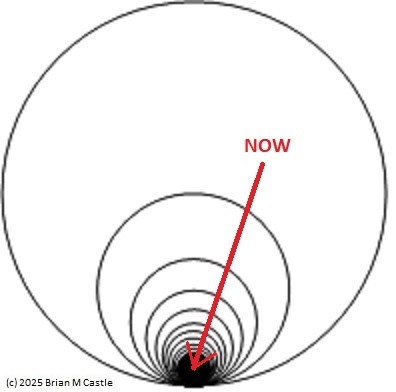 The earring has some interesting mathematical properties. For one, it is self similar; any subset of the base point is homeomorphic to the entire structure. And, the base point is the only point where the neighborhood fails to be simply connected. When the earring is continuous, it is a space filling curve, it is a one dimensional Peano continuum. 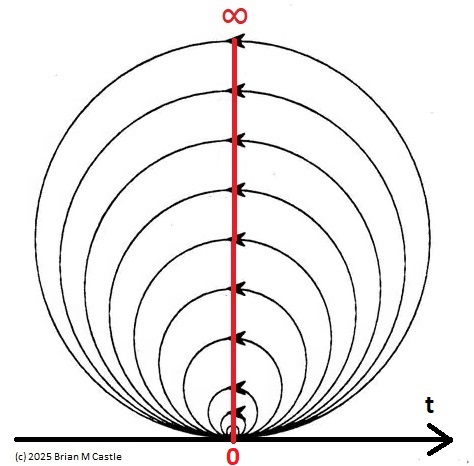 To emphasize, the radius of a loop models the length of a reflex arc. A reflex arc is viewed in the broadest possible terms, as a directional signal flow that can occur at any level of resolution. There are reflex arcs in the ordinary (anatomical and electrophysiological) sense, and there are also reflex arcs at the molecular level, and at the level of entire neural networks and subsystems. These loops have two things in common - they all intersect t=0 "now", and their points at infinity all line up in the compactified space. 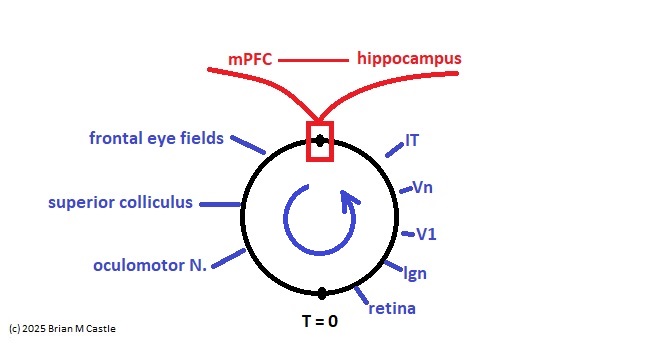 Note that the projected (compactified) view extends beyond the boundaries of the linear timeline (the green line, in the figure below - we got this when we added the point at infinity). However the resolution of the neighborhood around T=0 is considerably greater than that near the boundaries. The compactification has given us an almost "foveal" insight to the area we most need to decipher in a real time setting. And note that the neighborhood of infinity (the point labeled "observer" in the drawing) is precisely the area where we wish to place the hippocampus and prefrontal cortex, based on the earlier discussion of the stylized timeline.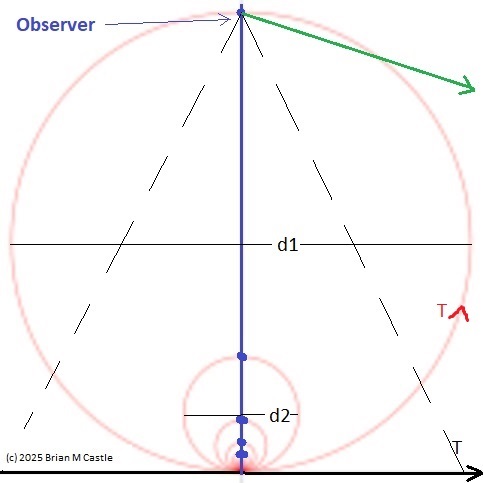 The neural timeline can be considered as an extension of a time series in which future points are predicted. Predictions become constrained (and therefore one hopes more accurate) as we move leftward along the timeline from T >> 0. In this sense each neural network layer along the timeline acts like a Kalman filter, predicting the next network state from the information within a moving window. This idea dovetails quite perfectly with concepts from machine learning like predictive coding and energy minimization. 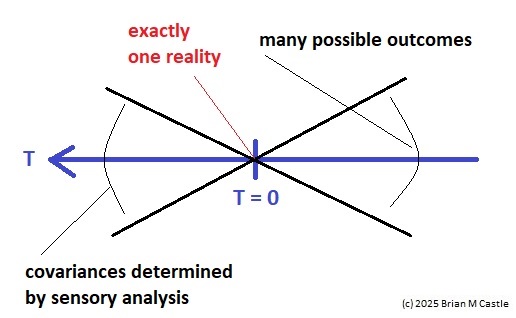 A complete mathematical description of the earring topology is beyond the scope of these pages. It is well studied, including its fundamental group and its embeddings (de Smit 1992, Eda and Kawamura 2000). (The information is readily available online, with a simple Google search). Here for example is an earring considered as a cross-section of a toroidal spiral. Note that in the description above, the origin is implicitly defined as the intersection of the circles that make up the earring. But we've already seen, that the origin is arbitrary - we can move it anywhere. With a simple rotation, we can move it to the opposite side of the circle, directly opposite the intersection of the hoops. After talking briefly about the ability to resolve individual points along the timeline, we'll see that with the proper embedding, we can even move the origin inside the circle. Embeddings are introduced on the next page.A Practical ApplicationSetting aside for a moment the issues around the neighborhood of infinity, here is a practical application of the compactified timeline (especially if you're a stock trader). We can do multi-window Granger causality analysis on any two compactified time series taken from the timeline, by simply rotating the circles. Here is a schematic of the underlying calculations. Look familiar?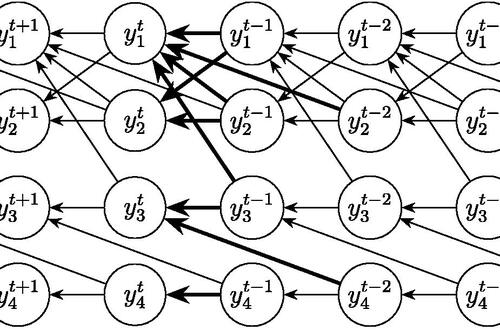 Causality analysis involves sliding the two time series relative to each other, which in the compact space becomes a rotation. When appropriately embedded, the output of the compactified network results in maps like this (and note that these are relationships in time, each arrow represents a correlation with a particular lag - for example if this were the stock market we might be interested in the one way arrow between livestock and supply shock, indicating both the relevant delay, and the corresponding correlation or likelihood).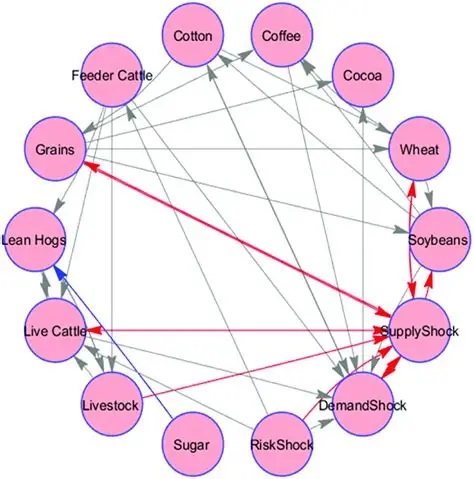 Instead of market data though, in our case the circles will attach to points along the timeline, and we'll be extracting "micro-causality" in real time from this network. We do not make the assumption that causality is stationary, instead we learn to identify and exploit the non-stationarities. If you're a stock trader, you can do this in real time with a neural network, you don't have to wait overnight for an analysis. You can take your nVidia GPU with you to the trading floor. ;) In a human brain, the idea of weighted connections along the timeline becomes closely related to real time abilities. In a biological context "real time" is mostly defined by the environment, in other words the organism is optimized to be effective in its environment, and it doesn't have to exceed that unless there is a specific need. However in the timeline model presented here, "real time" has a deeper meaning. The model is topologically self-similar, which means real time should extend all the way to the limit as dt => 0. This means "environmental speed" is no longer adequate, we need faster processes. If we want to get down to the range of molecular kinetics, we need at least microseconds - the milliseconds from a neural action potential aren't quite fast enough. Before expanding this model, it helps to have an idea of the range of time scales in play. Many scientists think of neural activity in terms of milliseconds, perhaps seconds, and this range is likely reasonable for single neurons. However neurons in populations acquire considerably better resolution, and this becomes vitally important in the consideration of asynchronous information processing architectures.Next: Timeline Resolution |