The spectrum of eye movements includes reflexes that draw the eyes to objects of interest (which could be called visually driven attention), and precise voluntary movements that can occur even in the absence of visual input, in the dark or when the eyes are closed. There is also saccadic suppression of visual input, somehow our brains know that a saccade is about to occur and suppress the blur and retinal slip that would ordinarily accompany it.
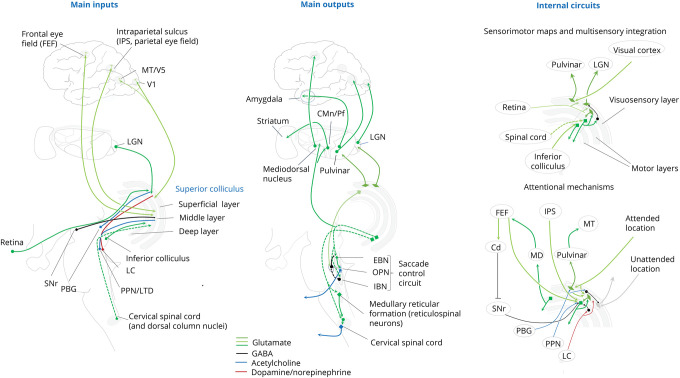
Most eye movements are ultimately under the control of the cerebral cortex. The three most relevant pathways are those from the frontal eye fields, parietal eye fields, and visual cortex to the superior colliculus. In turn, the SC projects back into each of these cortical areas, via the thalamus. The connections of greatest interest to us are those to and from the frontal eye fields (FEF) and the motion sensitive areas of the visual temporal lobe, areas MT and MST. These pathways serve different functions and are specialized anatomically.
The organization of the oculomotor system in the cerebral cortex can best be understood by examining its connections with the superior colliculus. In humans, the superficial layers of the SC are linked primarily with the visual cortex and posterior thalamus, receiving inputs from almost the entire visual cortex, area LIP in the parietal cortex, and the frontal eye fields. The intermediate layers connect with the auditory and somatosensory cortex and the posterior parietal cortex, and receive inputs from the parietal and frontal eye fields. The deep layers connect primarily with the brainstem and the cerebellum, and send corollary discharge information to the frontal eye fields. (Diano et al 2025)
Overall, the circuitry around the superior colliculus is as shown:
Superficial Layers of the Superior ColliculusThe superficial layers of the superior colliculus are visual, they receive direct retinal input and are mapped retinotopically.
The SC receives input from the early stages of visual processing, including the LGN, V1, and especially complex cells in V2. Additionally, there are massive projections from the frontal and parietal eye fields, and areas MT and MST of the temporal lobe, which process visual motion.
The SC receives input from the lateral intraparietal area (Gaymard et al 2003).
The SC receives input from the frontal eye fields (Sommer and Wurtz 2000).
The superficial SC projects back to area MT of the temporal lobe, as well as occipital and parietal cortex, via the inferior pulvinar nucleus PIm of the thalamus. This pathway is involved in saccadic suppression of visual input. There is also another small pathway from SC to the lateral shell region of the inferior pulvinar (Wurtz et al 2012). Unlike the pathway to the frontal eye fields (discussed below), these pathways do not carry corollary discharge information in advance of an eye movement. Instead, they handle saccadic suppression during the movement. Activity during saccades is visible both in the SC and in MT, as well as in the pulvinar. An example of saccadic suppression is show in the figure.
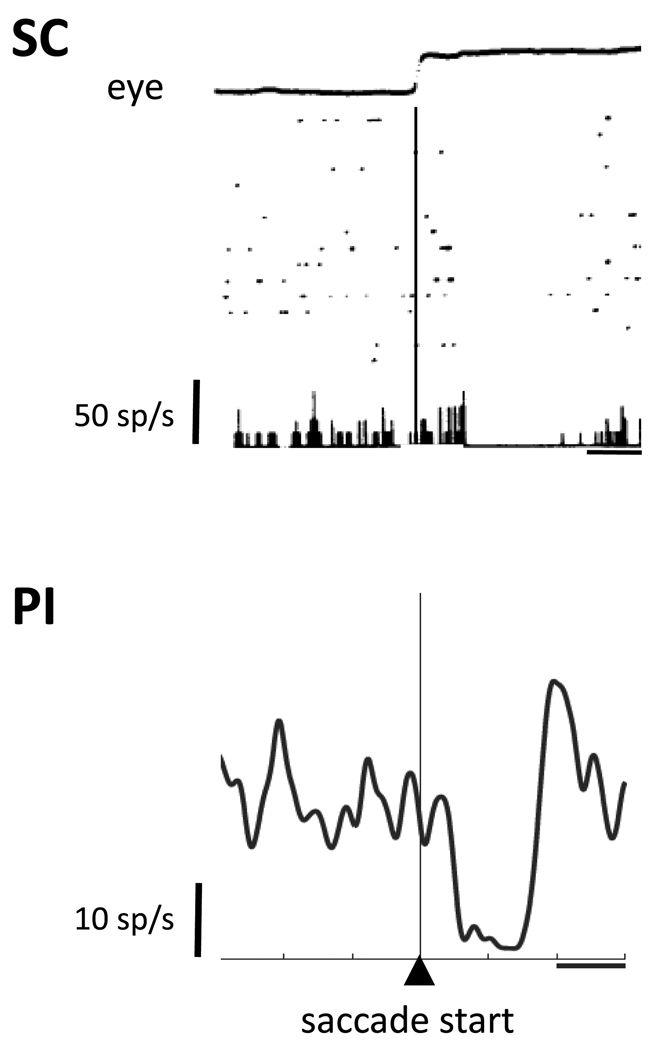
(figure from Wurtz et al 2012)
Another of the interesting features of saccades is a "gain control" for visual attention, related to the object (or portion of the visual field) that is being attended to - which in many cases corresponds to the target of a saccade. Neurons representing the attended stimulus fire more strongly, relative to their firing in relation to an unattended stimulus. This phenomenon appears to originate in the SC, and similar behavior is observed in its targets. The pathway for this gain control is still unknown, but it likely involves the dorsal medial region of the pulvinar, Pdm. In addition to changes in firing level, there are changes in latency (Maunsell 2021). Some of these are derived from stimulus intensity and are more closely associated with the retina, while others are derived from changes in attention and are more closely associated with the superior colliculus and thalamus.
Another important area of the thalamus called "thalamic reticular nucleus" (TRN) is involved in attentional control and modulation. This nucleus is basically a shell around the thalamus, and it receives excitatory inputs from both P and M cells in the LGN (which are thought to be collaterals of the LGN => V1 relay neurons), as well as the SC directly. TRN in turn, sends inhibitory projections back to both of the aforementioned layers of the LGN. TRN receives collaterals from the descending feedback neurons originating in layer 6 of V1, as well as ascending inputs from the superior colliculus. The superficial SC also targets the LGN directly. Apparently TRN does not target the superior colliculus directly.
Intermediate and Deep Layers of the Superior ColliculusThe intermediate and deep layers of the superior colliculus are mainly involved with saccade generation.
The intermediate SC projects back to the frontal eye fields via the parvocellular region of the medial dorsal nucleus of the thalamus (MD). This pathway is thought to mediate the "future field" effect in frontal eye field neurons, which is that the FEF neuron starts responding to visual input in the saccadic target location, before the saccade actually occurs. This pathway is a prime candidate for testing the timeline model. The "future field" effect is shown in the diagram.
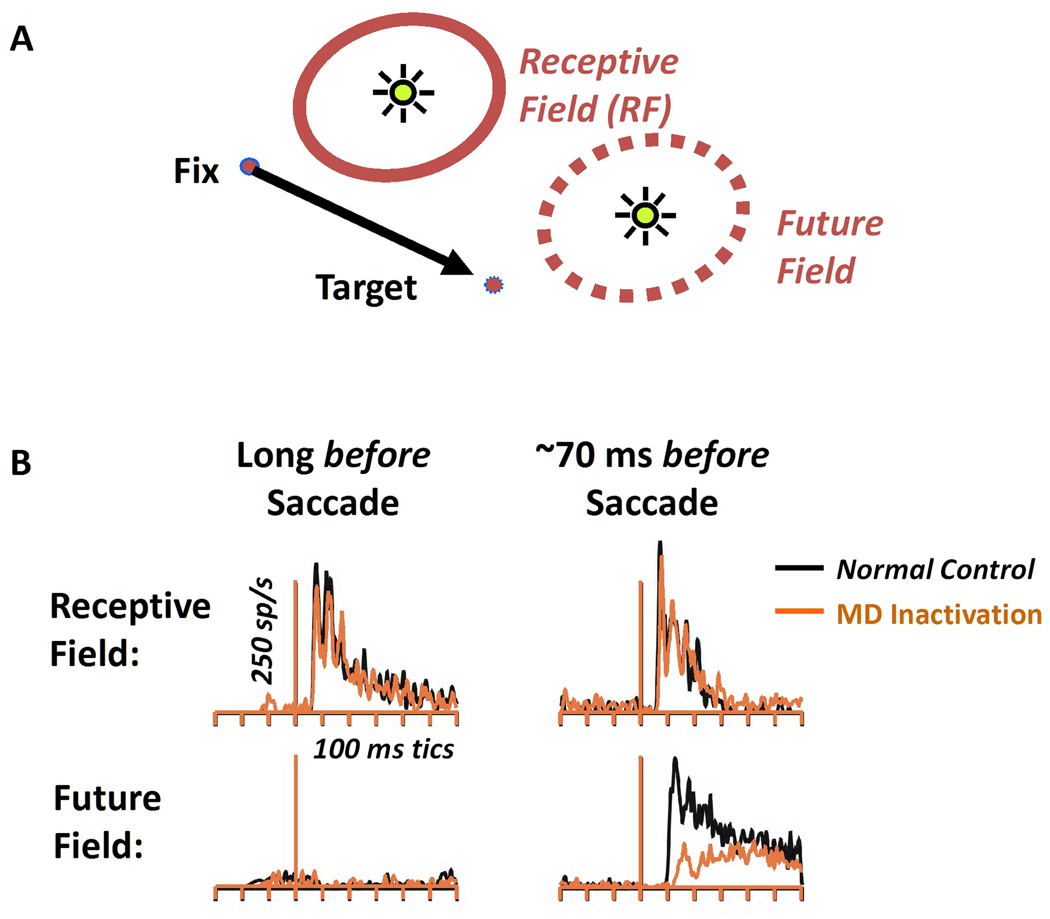
(figure from Wurtz et al 2012)
There is also a pathway from the intermediate SC to the lateral pulvinar nucleus of the thalamus PL, and area Pdm at the border of the lateral and medial pulvinar. These areas of the pulvinar in turn project to parietal and occipital cortex. This pathway carries preparatory information related to saccades (a "corollary discharge" when a saccade is about to occur).
In summary then, there are two major ascending pathways from the superior colliculus into the cerebral cortex, one from the superficial layers of the SC related mostly to visual attention and saccadic suppression, and another from the deeper layers of the SC carrying corollary discharge information to the frontal eye fields.
Control of Vergence Eye MovementsMost vergence movements are target driven. However it is possible for humans to emit disjunctive eye movements in the absence of visual input. The vergence system in the brain goes outside of the other oculomotor circuits, it is a separate set of pathways.
Next: Self Organization |